-
KIARAREBEL+2

 .
.User deleted
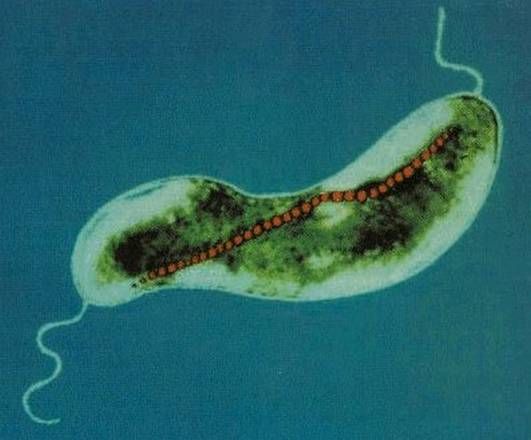
BATTERI MAGNETOTATTICI
I batteri magnetotattici sono una classe di batteri scoperta negli anni 1960 che presentano la particolarità di disporsi lungo le linee del campo magnetico terrestre (magnetotassi).
Introduzione
Il primo articolo su questa classe di batteri risale al 1963, ad opera di Salvatore Bellini, un laureato in medicina, interno presso l'Istituto di microbiologia dell'Università di Pavia. Egli infatti notò che sul suo vetrino da microscopia sul quale era depositato del fango di palude, un gruppo di batteri si muoveva decisamente in una direzione. Capì presto che questi microrganismi stavano seguendo le linee del campo magnetico terrestre, da sud a nord, da cui l'aggettivo "magnetosensibili". In seguito, nel 1975 un laureato in microbiologia, Richard Blakemore, pubblicò un articolo su "Science" e denominò i batteri con l'aggettivo "magnetotattici".
Questi batteri sono stati sottoposti ad innumerevoli esperimenti, sono persino stati portati a bordo dello Space Shuttle per esaminarne la magnetotassi in assenza di gravità, senza però giungere a conclusioni eclatanti. Sono anche stati "accusati di esistere" su Marte per via del ritrovamento di particelle magnetiche su un meteorite che si crede provenga dal pianeta rosso, anche questa volta l'incertezza dei risultati è perlomeno manifesta. Di certo invece è la singolarità di questi batteri, che probabilmente torneranno molto utili all'umanità in un futuro prossimo.
Biologia
I batteri magnetotattici (MagnetoTactic Bacteria, MTB) si trovano usualmente nella zona di transizione acqua-sedimenti (Oxic-Anoxic Transition Zone, OATZ) e ne esistono di diversa morfologia (bastoncellari, coccoidei, vibrioidi, spiraliformi). Diversi MTB contengono un differente numero, disposizione e forma di particelle magnetiche dei batteri (Bacterial Magnetic Particle, BMP). I MTB possono essere suddivisi in due categorie, a seconda se producono particelle di magnetite (IIIFe2IIFeO4) o di greigite (IIIFe2IIFeS4), anche se alcune specie le producono entrambe. La magnetite ha un momento magnetico tre volte maggiore della greigite.
La maggior parte degli MTB mineralizzanti magnetite necessitano di un ambiente microaerofilo per generare i magnetosomi. Sopra una certa soglia però non producono più BMP e quindi perdono la magnetotassi. Alcuni ceppi invece producono magnetite anche in condizioni anaerobiche, usando il monossido di azoto o lo ione nitrato come accettori finali di elettroni. Comunque fanno parte del sottogruppo alfa dei Proteobatteri. I MTB mineralizzanti grigite sono probabilmente stretti anaerobi e sono associati ai batteri solfato riduttori, quindi rientrano nel sottogruppo delta dei Proteobatteri. Questo indica un'origine evoluzionisticamente separata e quindi un processo di biomineralizzazione fondamentalmente diverso.
Si pensa che il vantaggio evolutivo dell'avere un sistema di magnetosomi consista nel riuscire a trovare e mantenere efficientemente una posizione ottimale in termini di sostanze chimiche e gradienti redox, riducendo una ricerca tridimensionale ad una monodimensionale (vedi prosieguo per il meccanismo). Recentemente si è appurato che alla magnetotassi è associato anche un meccanismo aerotattico.
Le BMP interagiscono formando delle catene. Nell'immagine se ne nota una in Magnetospirillum magneticum. Il dipolo magnetico della cellula è quindi la somma dei dipoli delle singole BMP ed è grande abbastanza da orientare passivamente la cellula e sopraffare le forze termiche casuali di un ambiente acquoso. In presenza di più di una catena, le forze di repulsione intercatena spingeranno le stesse ai bordi della cellula, inducendo turgore.
Magnetismo
Dal punto di vista fisico la crescita di un cristallo magnetico è governata da due fattori, uno tende ad allineare il momento magnetico delle molecole in soluzione con quello del cristallo in crescita, l'altro al contrario tende a ridurre il momento magnetico del cristallo permettendo l'attacco delle molecole in soluzione con un momento magnetico opposto. In natura ciò causa l'esistenza di domini magnetici, circondati da mura di dominio, dello spessore di circa 150 nm nella magnetite, nelle quali le molecole gradualmente cambiano orientazione. Per questo motivo macroscopicamente il ferro non è magnetico in assenza di un campo applicato. Similarmente particelle magnetiche molto piccole non presentano magnetizzazione a temperatura ambiente, il loro momento magnetico è alterato continuamente dai moti termici degli atomi che le compongono. Le MTB invece sono di dimensioni comprese tra 35 e 120 nm, cioè grandi abbastanza per avere un momento magnetico permanente ed allo stesso tempo piccole a sufficienza da essere un singolo dominio magnetico.
modello di polarità dei MTB
L'inclinazione del campo magnetico terrestre nei due emisferi seleziona una delle due possibili polarità delle cellule magnetotattiche (rispetto al polo flagellato della cellula), dirigendo la biomineralizzazione delle BMP. Infatti più del 99,9% della popolazione di MTB dell'emisfero nord si orienta e nuota verso il nord, quindi scendendo di livello (a causa della forma delle linee di campo) verso i sedimenti, allontanandosi da concentrazioni tossiche di ossigeno. Viceversa, ma con gli stessi risultati, nell'emisfero sud. All'equatore, dove le linee del campo geomagnetico sono orizzontali, si trovano all'incirca lo stesso numero di cellule con entrambe le polarità.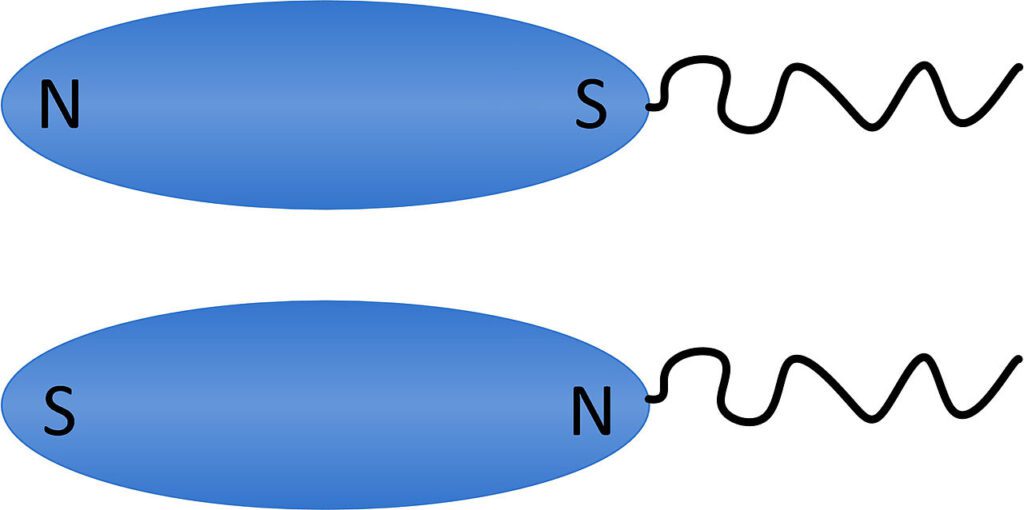
Magnetosoma
La biomineralizzazione della magnetite richiede dei meccanismi regolatori della concentrazione di ferro, della nucleazione del cristallo, del potenziale redox e del pH. La compartimentalizzazione in magnetosomi permette il controllo biochimico di tali processi. Dopo il sequenziamento dei genomi di alcune specie di MTB è stata possibile un'analisi comparativa delle proteine coinvolte nella formazione delle BMP. Si sono così trovate omologie di sequenza con membri dell'ubiquitaria famiglia CDF (Cation Diffusion Facilitator) e con delle serin proteasi Htr-simili. Le prime sono coinvolte esclusivamente nel trasporto di metalli pesanti, le seconde sono delle HSP che degradano le proteine mal ripiegate. Queste proteine della Membrana Magnetosomiale (MM) oltre al dominio serin proteasico contengono domini PDZ. Altre proteine della MM presentano domini TPR (Tetratrico Peptide Repeat).
TPR domain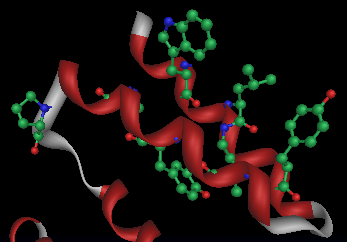
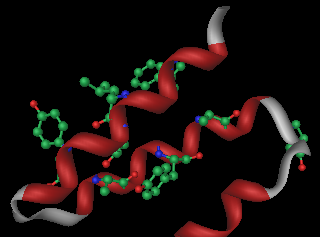
I domini TPR si ripiegano in due alfa-eliche e presentano una sequenza di consenso di 8 amminoacidi (dei 34 totali) molto conservati e diffusi in natura. A parte questi, il resto della struttura è specializzata a seconda delle implicazioni funzionali. I complessi noti che comprendono proteine TPR sono:
complessi di membrana che legano proteine e le trasportano all'interno di mitocondri e/o perossisomi
complessi che riconoscendo proteine leganti il DNA reprimono la trascrizione
l'APC (Anaphase Promoting Complex)
Esistono esempi sia di interazioni TPR-TPR che TPR-nonTPR[7] I modelli mostrano gli 8 residui conservati come sfere-bastoncini da due angolazioni diverse: W-LG-Y-A-F(qui Y)-A-P.
PDZ domain
I domini PDZ sono strutture modulari costituite da 6 beta-filamenti e 2 alfa-eliche che riconoscono gli amminoacidi C-terminali (almeno 4) delle proteine in modo sequenza specifico. Spesso il terzultimo residuo è fosforilabile, il che impedisce l'interazione col dominio PDZ. Gli unici residui conservati sono quelli deputati al riconoscimento del COOH terminale (RKXXXGLGF). Sono diffusi in natura in quanto costituiscono la struttura su cui si assemblano i complessi multiproteici, soprattutto quelli associati a proteine di membrana come i canali K+inward rectifier o i recettori beta2-adrenergici.
Membrana e proteine
La formazione della MM prevede almeno tre passaggi. Nel primo si forma un'invaginazione della membrana citoplasmatica, innescata da una GTPasi. È probabile che questo meccanismo sia simile anche negli eucarioti.
Il secondo passaggio prevede l'entrata di ioni ferrosi nelle vescicole neoformate a partire dagli ioni ferrici dell'ambiente esterno. Anche in colture Fe3+deficienti i MTB riescono ad accumularne elevate concentrazioni intracellulari secernendo un sideroforo, un ligando a basso peso molecolare con alta affinità e specificità per il Fe3+indotto al bisogno. Il complesso Ferro-sideroforo è quindi traslocato nel citoplasma, dove si scinde. Gli ioni ferrici devono quindi esser convertiti in ferrosi per poter essere accumulati nelle BMP ad opera di un trasportatore transmembrana con omologie di sequenza con antiporto Na+/H+. In realtà è un antiporto H+/Fe2+funzionante a gradiente protonico. Esso è localizzato sia sulla membrana citoplasmatica che sulla MM, ma in orientazioni invertite, in modo che sulla prima genera un efflusso di ferro e sulla seconda un influsso. Essendo decisamente più presente sulla MM, l'efflusso di ferro dalla membrana citoplasmatica è trascurabile. Inoltre questo step è controllato strettamente da un sistema redox di citocromi ancora non ben compreso e, sembra, specie-specifico.
Nello stage finale viene innescata la nucleazione del cristallo di magnetite ad opera di proteine transmembrana con domini idrofili acidi e basici. Una di queste proteine, chiamata Mms6, è stata usata nella sintesi artificiale di magnetite, e la sua presenza permette la produzione di cristalli omogenei in forma e dimensioni.
Probabilmente molte altre proteine associate alla MM svolgono ruoli nel generare concentrazioni di supersaturazione di Ferro, nel mantenere condizioni riducenti, nell'ossidazione del Ferro o nella parziale riduzione e deidratazione del ferridrato.
Biomineralizzazione
Nelle colture di Magnetospirillum magnetotacticum il ferro non è sostituibile da altri metalli di transizione (Ti, Cr, Co, Cu, Ni, Hg, Pb) eventualmente presenti nel terreno, anche se forse ciò è dovuto al trattamento. Allo stesso modo è stato visto che l'ossigeno e lo zolfo non sono intercambiabili come componente non metallica del magnetosoma in una stessa specie. Questo è indice dell'esistenza di diversi set di geni per la biomineralizzazione della magnetite e della grigite.
Dal punto di vista termodinamico la sintesi inorganica della magnetite è favorita rispetto a quella di altri ossidi del ferro a pH neutro ed a bassi potenziali redox. Sembra quindi che le condizioni microaerofile od anaerobie creino un potenziale adatto alla formazione delle BMP. Inoltre l'influsso di Ferro è rapidamente convertito in magnetite, senza ritardi, il che indica che la formazione dei cristalli non è preceduta da un accumulo di ferro sotto altre forme e che le strutture e gli enzimi per la biomineralizzazione sono già presenti nella cellula. Queste conclusioni sono supportate anche dal fatto che MTB coltivati in condizioni aerobiche (quindi non magnetici) contengono quantità di ferro paragonabili a qualsiasi altra specie batterica.
Applicazioni
Le caratteristiche delle BMP permettono la loro utilizzazione pratica in svariati campi, migliorando vecchie tecniche ed introducendone di nuove. Vantaggi comuni in tutti settori sono:
essendo circondate da un doppio strato lipidico formano aggregati facilmente disperdibili in ambiente acquoso, contrariamente alle particelle ottenute artificialmente.
le loro dimensioni le rendono superparamagnetiche, cioè seguono velocemente i cambi di campo magnetico esterno senza rimasugli delle polarizzazioni precedenti.
Biotecnologie
Negli ultimi anni l'uso di tecniche magnetiche è esploso e si è diversificato molto nel campo biotecnologico. I vantaggi principali sono l'alta scalabilità, rapidità e l'uso di reagenti non tossici ne pericolosi. La maggior parte delle particelle magnetiche artificiali (AMP) usate ricade in una di tre categorie:
particelle nude, ricoperte con polimeri (cellulosa, poliacroleina, silano, vetro borosilicato poroso, polistirene, resine a scambio ionico) derivati chimicamente per introdurre gruppi superficiali come COOH, NH2, alchilammine a lunga catena, prodotti dell'idrazide
particelle coniugate a ligandi con specificità generale, come streptavidina, oligo-dT, ProteinaA, ProteinaG
particelle coniugate a ligandi ad alta specificità, come Ig ed enzimi
Le dimensioni ricadono per lo più nel range 1-5 µm di diametro, rendendo necessario un continuo mescolamento per avere un efficiente binding. Di contro le forze di taglio possono danneggiare i ligandi, sia cellule che biomolecole, se si eccede nell'agitazione. Nel caso si abbia a che fare con popolazioni cellulari può essere necessario staccare le AMP dalle cellule, che risentirebbero della loro ingombrante presenza. Per questi motivi le BMP potrebbero trovare il loro spazio nella ricerca biotecnologica in sostituzione delle AMP, almeno in certe applicazioni.
Un'interessante integrazione delle particelle magnetiche con le tecniche di DNA ricombinante prevede l'uso di vettori esprimenti un marker di superficie come il CD4 o le MHC-I di topo. In tal modo è possibile, usando particelle magnetiche coniugate con un anticorpo contro tali marker, isolare in gran rapidità le cellule trasfettate con successo. Questa tecnica potrebbe essere estesa a cellule procariote, e l'uso di BMP ricombinanti esprimenti l'Ig di selezione potrebbe rendere indipendente un laboratorio di ricerca dall'acquisto di sempre nuove AMP coniugate con l'anticorpo.
Le BMP si prestano molto bene come carrier di proteine ricombinanti o di proteine ancora, con applicazioni nella ricerca, nella diagnostica e come biosensori. In un esperimento classico, usando una proteina di fusione MagA-Luc (Luc=Luciferasi, MagA=proteina dei MTB), si è dimostrato che MagA è localizzata sulla superficie delle BMP. Quindi con un vettore di espressione plasmidico si è fatta esprimere in un MTB la proteina ricombinante MagA-ProteinaA, che grazie alla parte derivata da MagA si localizza sulle BMP e per mezzo della ProteinaA può legare la parte costante delle Ig. Isolate le BMP ricombinanti le si è incubate con l'anticorpo anti-IgG coniugato con la Fosfatasi Alcalina. In questo modo si è potuto dimostrare, usando un substrato luminescente per la Fosfatasi Alcalina, che l'anticorpo era effettivamente legato alle BMP perché la ProteinaA era fusa con MagA. Contrariamente, BMP prodotte da MTB esprimenti MagA e ProteinaA separate emettevano una luminescenza trascurabile, segno che la ProteinaA da sola non ha affinità per le BMP.
Per aumentare il segnale luminoso di questo saggio occorre aumentare la quantità di ProteinaA espressa sulle BMP e quindi usare un vettore di espressione più forte, che sia cioè ad alto numero di copie, stabile e derivato dall'ospite. In Magnetospirillum magneticum è stato scoperto un plasmide criptico denominato pMGT con queste caratteristiche. È lungo circa 3,7 kb ed ha due potenziali ORF codificanti le proteine Rep e Mob. Tagliando pMGT in modo che contenga la regione replicativa si ottiene un frammento di 3 kb funzionale. Nonostante elettroporazione (usata per trasformare il plasmide) uccida i MTB contenenti BMP, è possibile effettuarla in condizioni aerobiche, nelle quali le BMP non sono prodotte. Successivamente il ripristino dell'anaerobiosi permette ai MTB di riprendere a biomineralizzare le BMP.
Con caratteristiche simili il plasmide può essere impiegato per ricombinare a proteine della MM vari target della ricerca farmacologica: un esempio su tutti, i recettori a sette domini transmembrana. Proprio uno di essi è stato clonato ed espresso sulle BMP per dimostrare che la proteina ricombinante manteneva la caratteristica di legare i propri agonisti ed antagonisti (marcati con una molecola fluorescente). Questo sistema inoltre è in grado di quantificare l'interazione e può essere scalato per una larga produzione di BMP ricombinanti per Ig, enzimi, .. mantenendone l'attività.
Replicazione plasmidica
La minima regione replicativa deve contenere una sequenza di origine ori (ricca di AT, dove avviene la separazione delle eliche), delle sequenze ripetute per il legame di Rep (iteroni), i boxes per la DnaA (codificata dall'ospite) ed il gene rep. Il legame di Rep e DnaA favoriscono la distorsione dell'elica a livello di ori che, per l'alta percentuale di AT, si separa in due emieliche. Da qui parte la replicazione col legame delle proteine SSB e della DnaB (elicasi).
Medicina
Le caratteristiche delle BMP possono tornare molto utili in campo medico, in diagnosi come in terapia. Le AMP essendo prodotte per sintesi chimica o per via meccanica risentono delle loro disomogenee qualità fisico-chimiche. Inoltre le proprietà delle particelle dovrebbero essere riproducibili. Le possibili applicazioni mediche richiedono particelle superparamagnetiche come le BMP. Ovviamente occorre stabilità alle condizioni fisiologiche di pH, salinità ed osmolarità. Devono essere facilmente disperdibili e non tossiche. Quindi a seconda se queste particelle verranno usate in vivo od in vitro ci saranno altre restrizioni. Per applicazioni in vivo è necessario evitare la formazione di agglomerati, con rischio di embolia vasale. Qui tornano utili le dimensioni nanoscopiche delle BMP, che conferiscono bassissime velocità di sedimentazione (quindi stabilità colloidale), migliore diffusione tissutale ed enorme area superficiale. Quest'ultima proprietà è importante se si intende fissare sulla superficie delle BMP un ligando. La magnetite è tuttora il materiale magnetico più usato in medicina per la sua bassa tossicità ed immunogenicità. Per applicazioni in vitro le restrizioni sono minori.
Le applicazioni in vivo possono essere ulteriormente suddivise in terapeutiche e diagnostiche, mentre quelle in vitro sono principalmente diagnostiche per il momento. Fra le prime, le due tecniche più discusse sono l'ipertermia e la veicolazione di farmaci. Con la prima si intende una procedura terapeutica usata per innalzare la temperatura di una regione del corpo colpita da un tumore. È praticata in concomitanza con altre terapie anticancro e si basa sul fatto che sopra i 40-43 °C le cellule muoiono per shock termico. Le particelle magnetiche, dopo una somministrazione endovenosa, possono essere tenute in loco da un campo magnetico esterno, costante prima, ed alternato poi, causando un "effetto Joule" magnetico dovuto alla continua riorientazione della magnetizzazione delle particelle (che hanno bassa conducibilità elettrica). Le nanoparticelle assorbono molta più potenza in campi magnetici AC tollerabili e la loro uniformità dimensionale è fondamentale per un controllo preciso della temperatura.
Le stesse particelle possono essere usate per trasportare farmaci, radionuclei od anticorpi, per un rilascio controllato nel tempo e nello spazio, aumentando le concentrazioni in situ e diminuendo gli effetti collaterali. Nel caso di un tumore per esempio è possibile far extravasare le particelle magnetiche grazie ad un forte campo magnetico. Queste rimangono intrappolate nel tessuto maligno dove rilasciano il principio attivo o le radiazioni. Con le BMP è possibile preparare magnetoliposomi contenenti agenti antitumorali.
La risonanza magnetica nucleare ad immagine (iNMR) utilizza una classe particolare di farmaci, i cosiddetti magnetofarmaceutici, da somministrare al paziente per invigorire il contrasto tra tessuti sani e malati, per visualizzare la circolazione sanguigna e lo stato degli organi. Si possono così distinguere infarti cerebrali e cardiaci, lesioni epatiche, tumori, infiammazioni ed ischemie. Ancora una volta le piccole dimensioni fan risaltare differenze utili tra i tessuti come la composizione ed i processi endocitotici.
Nelle tecniche in vitro è possibile separare una popolazione cellulare che esprime un particolare epitopo in mezzo a molti altri tipi cellulari. Per lo scopo si sfrutta una selezione positiva se l'anticorpo è specifico per il tipo cellulare voluto, negativa (o di arricchimento) se gli anticorpi sono specifici per le cellule non-target.
Recentemente è stata introdotta una nuova tecnica di valutazione dei saggi immunologici chiamata magnetorilassometria. Essa misura la viscosità magnetica, cioè il rilassamento del momento magnetico netto, di un sistema dopo la rimozione del campo magnetico esterno. La viscosità magnetica dipende dalle dimensioni idrodinamiche e del nocciolo, e dall'anisotropia delle particelle, e permette di distinguere tra quelle coniugate con un ligando e quelle libere grazie al diverso comportamento magnetico.
VOCI CORRELATE:
GREIGITE
NANOROBOT
**************************************************************************************************************
www.treccani.it/enciclopedia/magne...nze-Fisiche%29/
MAGNETOTASSIA
Magnetotassìa (o magnetotassi o magnetotattismo) [Comp. di magneto- e tassia (o -tassi o tattismo)] Meccanismo con cui un organismo assume e mantiene una posizione orientata in relazione a un campo magnetico.
***************************************************************************************************************
BANCA DATI FAGO M-13 QUI : https://killdogma.forumfree.it/?t=78646181
Edited by Kill Dogma Revolution ! - 16/11/2021, 20:56 .
. -
KIARAREBEL .
User deleted
Swarm of Bacteria Builds Tiny Pyramid - LO SCIAME DI BATTERI COSTRUISCE UNA PICCOLA PIRAMIDE
. -
KIARAREBEL .
User deleted
https://ingvambiente.com/2019/09/17/hard-d...llultrapiccolo/
Hard disk, ferrofluido e batteri magnetotattici: il magnetismo dell’ultrapiccolo
Nel paleomagnetismo, come in tante applicazioni tecnologiche che utilizzano il magnetismo, è essenziale individuare minerali e materiali capaci di magnetizzarsi fedelmente al campo magnetico applicato, mantenendo tale magnetizzazione stabile nel tempo. Vediamone le caratteristiche e le peculiarità.
In natura, alcuni minerali di dimensione submicroscopica mostrano proprietà magnetiche particolari. Queste particelle, solitamente costituite da ossidi di ferro quali la magnetite (Fe3O4), sono al centro di molte ricerche a causa del loro possibile impiego in biomedicina, nell’imaging a risonanza magnetica, nell’immagazzinamento dati e nelle geoscienze. In questa rassegna si vogliono introdurre i principi che regolano la magnetizzazione dei micro e nanoparticolati, da cui derivano le rilevanti applicazioni tecnologiche.
I corpi capaci di acquisire una magnetizzazione stabile nel tempo si dicono ferromagnetici; essi sono costituiti da volumi elementari, detti domini, dotati di magnetizzazione propria. Sono quei materiali in grado di magnetizzarsi molto intensamente sotto l’azione di un campo magnetico esterno e di restare a lungo magnetizzati quando il campo si annulla, diventando così dei magneti. Vengono detti granuli singolo dominio (SD) quei minerali magnetici le cui dimensioni sono inferiori a quelle critiche per la separazione in più domini. Per un granulo di magnetite equidimensionale, tale valore teorico è pari a circa 0.080 m. Quando le dimensioni del granulo sono troppo piccole per costituire un dominio magnetico perché l’energia termica prevale sulla possibilità di mantenere una magnetizzazione rimanente, si parla di particelle superparamagnetiche (SP). Nel caso della magnetite, questo limite è circa 0.035 m. Pertanto, teoricamente, si parla di magnetite SD quando le dimensioni granulari sono comprese tra 0.035 e 0.080 m.
Nelle scienze della Terra, i minerali ferromagnetici permettono di studiare il campo magnetico del passato “fossilizzato” nelle rocce, su cui si basa la disciplina del paleomagnetismo. I minerali con proprietà magnetiche intense e stabili sono dimensionalmente SD e vengono utilizzati anche da chi costruisce dispositivi (per esempio hard disk) che debbano mantenere il loro stato di magnetizzazione nella maniera più stabile possibile nel tempo. Per fare un esempio, un granulo SD non equidimensionale di magnetite, di lunghezza 0.1 m e larghezza 0.02 m, a temperatura ambiente, perde la sua magnetizzazione in un tempo che supera l’età della Terra.
Il funzionamento di un hard disk dipende dall’associazione di ognuno dei suoi bit di informazione (1 o 0) a uno stato di magnetizzazione. La sua densità d’informazione (bit al pollice quadro) è data dal numero di domini che costituiscono un singolo bit, moltiplicato per la loro estensione superficiale media, diviso per la superficie di archiviazione. Quindi, per aumentare la quantità di dati sullo stesso disco, si deve ridurre il numero di domini che definiscono il singolo bit, oppure ridurre l’area disponibile per un singolo dominio magnetico. Nel caso in cui il numero di domini che definiscono un singolo bit si riduca all’unità e la loro area sia dell’ordine di pochi nm2, il comportamento diventa superparamagnetico, e le informazioni registrate vengono immediatamente perdute. Per questo, a parità di dimensione dell’hard disk determinata da standard costruttivi, esiste un limite fisico alla sua capienza.
In campo biologico, esiste una classe di batteri, denominati magnetotattici, che hanno la capacità di orientarsi secondo le linee di forza del campo magnetico (magnetotassi), contenendo i magnetosomi, particelle magnetiche SD rivestite da una membrana lipidica da loro stessi sintetizzate. I batteri usano il campo geomagnetico per orientarsi verso acque profonde ipossiche, a bassa concentrazione di ossigeno. Come si può studiare grazie a sofisticate tecniche magnetiche di laboratorio, i magnetosomi sono allineati in catene ordinate a debole interazione, in modo da impedirne l’agglomerazione. Il dipolo magnetico totale del batterio è sufficientemente intenso da permettere al batterio di orientarsi, vincendo i gradienti termici e le correnti agenti nell’ambiente acquoso. Questi batteri, scoperti nel 1963 da Salvatore Bellini, sono stati sottoposti a innumerevoli esperimenti, effettuati persino a bordo dello Space Shuttle, per esaminarne le caratteristiche in assenza di gravità. In questo video, si può osservare l’effetto di un magnete sul verso del moto di batteri magnetotattici. L’INGV ha recentemente collaborato all’identificazione, in prossimità della foce del fiume Neponset, in Massachussetts, di un batterio magnetotattico con una struttura a catena di cristalli di magnetite pura, ciascuno della dimensione di 20-50 nm.
I batteri magnetotattici hanno suggerito notevoli applicazioni nei settori biomedici e nanotecnologici; in questo link, si riportano i trattamenti oncologici innovativi effettuati da Sylvain Martel del Politecnico di Montreal, utilizzando il batterio Magnetococcus marinus (MC-1). Il meccanismo magnetotattico prevede l’applicazione di un debole campo magnetico per veicolare i batteri, caricati con un farmaco, in direzione del tumore. Quando i batteri raggiungono l’area tumorale, ipossica, rilasciano il farmaco, che agisce così in maniera localizzata su un’area specifica. A tale proposito, si rimanda a questo articolo per una rassegna scientifica sull’utilizzo dei batteri magnetotattici nella terapia tumorale.
Introduciamo adesso il ferrofluido, un olio contenente particelle magnetiche ultrafini superparamagnetiche, solitamente di dimensione intorno a 10 nm, rivestite di un tensioattivo per prevenirne l’agglomerazione. Il comportamento di un ferrofluido in presenza di un campo magnetico è peculiare: le particelle tendono ad allinearsi alle linee di forza del campo magnetico agente, generando curiose forme appuntite, simili a ricci.
I ferrofluidi, per la loro eccezionale risposta ai campi magnetici, sono impiegati in tutti quei dispositivi acustici e ottici in cui è essenziale una dinamica ottimale. A proposito di particelle SP, si noti che una delle loro applicazioni più comuni sono i toner, in cui gli ossidi di ferro che li compongono vengono veicolati e disposti magneticamente nel foglio di carta, per poi essere successivamente fusi, attaccandosi alle sue fibre.. -
KIARAREBEL .
User deleted
www.blog-erboristeriaweb.it/la-pro...-comportamento/
LA PROTEINA “MAGNETO” GENETICAMENTE MODIFICATA CONTROLLA A DISTANZA IL CERVELLO E IL COMPUTER
La proteina “Magneto” geneticamente modificata controlla a distanza il cervello e il comportamento.
La proteina “Magneto” geneticamente modificata controlla a distanza il cervello e il comportamento
La camera magnetica toroidale (Tokamak) del Joint European Torus (JET) presso il Culham Science Center. Fotografia: AFP / Getty Images
Il nuovo metodo “Badass” utilizza una proteina magnetizzata per attivare le cellule cerebrali in modo rapido, reversibile e non invasivo
I ricercatori negli Stati Uniti hanno sviluppato un nuovo metodo per controllare i circuiti cerebrali associati a comportamenti animali complessi, utilizzando l’ingegneria genetica per creare una proteina magnetizzata che attiva gruppi specifici di cellule nervose a distanza.
Capire come il cervello genera il comportamento è uno degli obiettivi finali della neuroscienza e una delle sue domande più difficili. Negli ultimi anni, i ricercatori hanno sviluppato una serie di metodi che consentono loro di controllare a distanza specifici gruppi di neuroni e di sondare il funzionamento dei circuiti neuronali.
Il più potente di questi è un metodo chiamato optogenetica , che consente ai ricercatori di attivare o disattivare popolazioni di neuroni correlati su una scala temporale millisecondo per millisecondo con impulsi di luce laser. Un altro metodo sviluppato di recente, chiamato chemogenetica , utilizza proteine ingegnerizzate che vengono attivate da farmaci di design e possono essere mirate a specifici tipi di cellule.
Sebbene potenti, entrambi questi metodi presentano degli svantaggi. L’optogenetica è invasiva, richiede l’inserimento di fibre ottiche che trasportano gli impulsi luminosi nel cervello e, inoltre, la misura in cui la luce penetra nel tessuto cerebrale denso è fortemente limitata. Gli approcci chemogenetici superano entrambi questi limiti, ma in genere inducono reazioni biochimiche che impiegano diversi secondi per attivare le cellule nervose.
Controllo remoto dell’attività cerebrale con nanoparticelle riscaldate
Leggi di più
La nuova tecnica, sviluppata nel laboratorio di Ali Güler presso l’Università della Virginia a Charlottesville e descritta in una pubblicazione online anticipata sulla rivista Nature Neuroscience , non solo non è invasiva, ma può anche attivare i neuroni in modo rapido e reversibile.
Diversi studi precedenti hanno dimostrato che le proteine delle cellule nervose attivate dal calore e dalla pressione meccanica possono essere ingegnerizzate geneticamente in modo che diventino sensibili alle onde radio e ai campi magnetici , legandole a una proteina che immagazzina il ferro chiamata ferritina oa particelle paramagnetiche inorganiche . Questi metodi rappresentano un progresso importante – ad esempio, sono già stati utilizzati per regolare i livelli di glucosio nel sangue nei topi – ma coinvolgono più componenti che devono essere introdotti separatamente.
La nuova tecnica si basa su questo lavoro precedente, e si basa su una proteina chiamata TRPV4, che è sensibile sia temperatura e forze traenti . Questi stimoli aprono il suo poro centrale, consentendo alla corrente elettrica di fluire attraverso la membrana cellulare; questo evoca impulsi nervosi che viaggiano nel midollo spinale e poi fino al cervello.
Güler ei suoi colleghi hanno ragionato che le forze di coppia magnetica (o rotanti) potrebbero attivare TRPV4 aprendone il poro centrale, e quindi hanno usato l’ingegneria genetica per fondere la proteina nella regione paramagnetica della ferritina, insieme a brevi sequenze di DNA che segnalano alle cellule di trasportare proteine alla membrana delle cellule nervose e inserirle in essa.
Manipolazione in vivo del comportamento del pesce zebra utilizzando Magneto. Le larve di pesce zebra mostrano un comportamento di avvolgimento in risposta a campi magnetici localizzati. Da Wheeler et al (2016).
Quando hanno introdotto questo costrutto genetico nelle cellule renali embrionali umane che crescono nelle piastre di Petri, le cellule hanno sintetizzato la proteina “Magneto” e l’hanno inserita nella loro membrana. L’applicazione di un campo magnetico ha attivato la proteina TRPV1 ingegnerizzata, come evidenziato da aumenti transitori della concentrazione di ioni calcio all’interno delle cellule, rilevati con un microscopio a fluorescenza.
Successivamente, i ricercatori hanno inserito la sequenza Magneto DNA nel genoma di un virus, insieme al gene che codifica per la proteina fluorescente verde e sequenze di DNA regolatorie che fanno sì che il costrutto venga espresso solo in tipi specifici di neuroni. Hanno quindi iniettato il virus nel cervello dei topi, mirando alla corteccia entorinale e sezionato il cervello degli animali per identificare le cellule che emettevano fluorescenza verde. Utilizzando microelettrodi, hanno poi dimostrato che l’applicazione di un campo magnetico alle fettine di cervello attiva Magneto in modo che le cellule producano impulsi nervosi.
Per determinare se Magneto può essere utilizzato per manipolare l’attività neuronale in animali vivi, hanno iniettato Magneto nelle larve di pesce zebra, prendendo di mira i neuroni nel tronco e nella coda che normalmente controllano una risposta di fuga. Hanno quindi posizionato le larve di pesce zebra in un acquario magnetizzato appositamente costruito e hanno scoperto che l’esposizione a un campo magnetico ha indotto manovre di avvolgimento simili a quelle che si verificano durante la risposta di fuga. (Questo esperimento ha coinvolto un totale di nove larve di pesce zebra e le analisi successive hanno rivelato che ciascuna larva conteneva circa 5 neuroni che esprimevano Magneto.)
I ricercatori leggono e scrivono l’attività cerebrale con la luce
Leggi di più
In un esperimento finale, i ricercatori hanno iniettato Magneto nello striato di topi che si comportano liberamente, una struttura cerebrale profonda contenente neuroni produttori di dopamina che sono coinvolti nella ricompensa e nella motivazione, e poi hanno posizionato gli animali in un apparato diviso in sezioni magnetizzate e non magnetizzate. . I topi che esprimevano Magneto trascorrevano molto più tempo nelle aree magnetizzate rispetto ai topi che non lo facevano, perché l’attivazione della proteina ha indotto i neuroni striatali che la esprimevano a rilasciare dopamina, in modo che i topi trovassero gratificante in quelle aree. Ciò dimostra che Magneto può controllare a distanza l’attivazione dei neuroni nel profondo del cervello e anche controllare comportamenti complessi.
Il neuroscienziato Steve Ramirez dell’Università di Harvard, che usa l’optogenetica per manipolare i ricordi nel cervello dei topi, dice che lo studio è ” tosto “.
“I tentativi precedenti [utilizzando magneti per controllare l’attività neuronale] richiedevano più componenti per il funzionamento del sistema: iniettare particelle magnetiche, iniettare un virus che esprime un canale sensibile al calore, [o] fissare la testa dell’animale in modo che una bobina potesse indurre cambiamenti nel magnetismo “, spiega. “Il problema con un sistema multicomponente è che c’è così tanto spazio per la scomposizione di ogni singolo pezzo.”
“Questo sistema è un singolo, elegante virus che può essere iniettato ovunque nel cervello, il che rende tecnicamente più facile e meno probabile che si rompano le campane e i fischietti in movimento”, aggiunge, “e la loro attrezzatura comportamentale è stata progettata in modo intelligente per contenere magneti dove appropriato in modo che gli animali possano muoversi liberamente. ”
La “magnetogenetica” è quindi un’importante aggiunta alla cassetta degli attrezzi dei neuroscienziati, che senza dubbio sarà ulteriormente sviluppata e fornirà ai ricercatori nuovi modi di studiare lo sviluppo e la funzione del cervello.
Riferimento
Wheeler, MA, et al . (2016). Controllo magnetico mirato geneticamente del sistema nervoso. Nat. Neurosci ., DOI: 10.1038 / nn.4265 [ Abstract ]
… poiché è il nostro mese di compleanno, abbiamo un piccolo favore da chiederti. Non capita tutti i giorni di compiere 200 anni. Dato che oggi vi raggiungete dall’Italia, speriamo che ci aiutiate a celebrare due secoli di giornalismo indipendente e ad alto impatto del Guardian. Dal 1821, decine di milioni hanno riposto la loro fiducia nella nostra reportistica di qualità, rivolgendosi a noi nei momenti di crisi, incertezza, solidarietà e speranza. E questo è solo l’inizio.
Grazie ai finanziamenti di oltre 1,5 milioni di sostenitori del Guardian in 180 paesi, siamo rimasti aperti a tutti e ferocemente indipendenti. Senza azionisti o proprietari miliardari, stabiliamo la nostra agenda e forniamo giornalismo alla ricerca della verità che sia libero da influenze commerciali e politiche. Quando non è mai più importante, possiamo indagare e sfidare senza paura o favore.
A differenza di molti altri, il giornalismo del Guardian può essere letto da tutti, indipendentemente da ciò che possono permettersi di pagare. Lo facciamo perché crediamo nell’uguaglianza delle informazioni. Un numero maggiore di persone può tenere traccia degli eventi globali, comprendere il loro impatto su persone e comunità e trarre ispirazione per intraprendere azioni significative.
Negli ultimi 200 anni, abbiamo riportato le questioni e gli eventi più critici della giornata e pubblicato innumerevoli esclusive che hanno ritenuto i potenti responsabili ed esposto incompetenza, ingiustizia e scorrettezza. Da Windrush a Wikileaks, da Snowden a Cambridge Analytica, dalla crisi climatica alla discriminazione razziale, i nostri giornalisti indagano sulle azioni illecite nella speranza che rivelarle possa aiutare a rimediare.
Se c’è mai stato un momento per unirsi a noi, è adesso. Ogni contributo, piccolo o grande che sia, alimenta il nostro giornalismo e sostiene il nostro futuro.
Edited by KIARAREBEL - 17/11/2021, 03:27.

 Kill Dogma Tv-Nella Tana del Bianconiglio-Illuminati&Nuovo Ordine Mondiale NWO-Il manuale del risveglio-Ricerca-Indagini-Notizie
Kill Dogma Tv-Nella Tana del Bianconiglio-Illuminati&Nuovo Ordine Mondiale NWO-Il manuale del risveglio-Ricerca-Indagini-Notizie  BANCHE DATI NO ALLA SPERIMENTAZIONE CLANDESTINA-Fonti&Prove:Vaccini,Microchip,Transumanesimo,5G,Mondo Smart City Intelligenti
BANCHE DATI NO ALLA SPERIMENTAZIONE CLANDESTINA-Fonti&Prove:Vaccini,Microchip,Transumanesimo,5G,Mondo Smart City Intelligenti -
 ">
">
BATTERI MAGNETOTATTICI |